Frühe Versuche
Wie lange dauert es eigentlich, bis ein Aktionspotenzial vom Axonhügel bis zur Synapse gelangt? Mit dieser interessanten Frage haben sich Biologen schon sehr früh beschäftig, und vor allem Hermann von HELMHOLTZ (1821 - 1894) ist hier zu nennen. Ende des 19. Jahrhunderts konstruierte er eine Versuchsanordnung, mit der er die Geschwindigkeit der Erregungsleitung ziemlich genau bestimmen konnte. Er isolierte dazu einen Froschmuskel samt zuleitendem motorischen Nerv. Diesen Nerv reizte HELMHOLTZ elektrisch und maß dann die Zeit, die bis zum Zusammenzucken des Muskels verging. Wenn er den Nerv ziemlich dicht am Muskel reizte, kontrahierte dieser schon nach wenigen Millisekunden. Reizte er den Nerv 3 cm weiter entfernt vom Muskel, dauert es ca. 1/100 Sekunde länger, bis dieser kontrahierte. Natürlich hatte HELMHOLTZ keine Uhr, mit der er derart genau die Zeit messen konnte. Er befestigte eine Art Stift an dem Muskel, und dieser Stift zeichnete eine Linie auf einer mit Kohle beschichteten Walze, die sich schnell drehte. Sobald sich der Muskel zusammenzog, ging der Stift nach oben und gleich wieder nach unten, und auf der Walze entstand so eine weiße Kurve.
Aus seinen Experimenten schloss HELMHOLTZ, dass sich die Erregung am Nerven mit ca. 3 m/s fortpflanzt. Schauen wir uns nun an, was die aktuelle Wissenschaft über die Erregungsleitungsgeschwindigkeit herausgefunden hat:
| Tierart | Durchmesser in mm | Geschwindigkeit in m/s |
| Qualle | 0,009 | 0,5 |
| Schabe | 0,050 | 7 |
| Tintenfisch | 0,650 | 25 |
Wenn man bedenkt, welche bescheidenen Mittel HELMHOLTZ im vorletzten Jahrhunder zur Verfügung standen, ist sein Ergebnis von 3 m/s schon ziemlich beeindruckend. Die Tabelle zeigt uns, das die Geschwindigkeit der Erregungsleitung sehr stark vom Durchmesser des Axons abhängt. Je dicker das Axon, desto schneller wird die Erregung weitergeleitet. Das liegt an der zunehmenden Oberfläche des Axons, die ja mit dem Radius linear ansteigt (Umfang eines Kreises = Pi * Radius).
Geschwindigkeiten bei Wirbeltieren
Bei Säugetieren wie der Katze hat man aber Geschwindigkeiten von 85 m/s und mehr gemessen. Welchen Durchmesser muss dann ein Axon der Katze haben, damit diese Geschwindigkeit erreicht wird?
Solche Fragen kann man mit Hilfe mathematischer Software leicht beantworten:
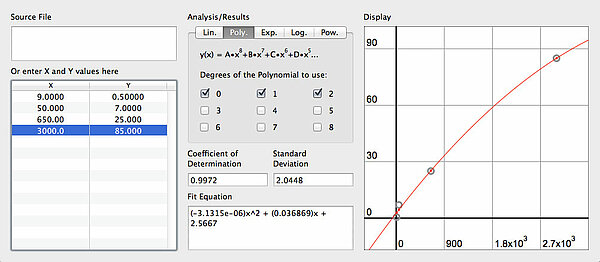
Ich habe hier in das Program LSAnalyzer einfach mal die Daten aus der obigen Tabelle eingetragen und für die Katze einen Durchmesser von 3 mm gewählt. Die quadratische Funktion passt sehr gut zu den vier Wertepaaren.
Wenn die Leitungsgeschwindigkeit allerdings nur von der Oberfläche des Axons abhängig wäre, müsste sie linear mit dem Radius ansteigen, was hier aber nicht der Fall ist. Offensichtlich spielen auch andere Faktoren bei der Leitungsgeschwindigkeit eine wichtige Rolle.
Ein Axon mit einem Durchmesser von 3 mm ist schon sehr dick, aber theoretisch noch denkbar. Ein Nerv allerdings besteht aber aus einem Bündel vieler solcher Axone und hätte damit leicht einen Durchmesser von 10 cm oder mehr, was bei einem kleinen Tier wie der Katze undenkbar ist.
| Tierart | Durchmesser in mm | Geschwindigkeit in m/s |
| Tintenfisch | 0,650 | 25 |
| Frosch | 0,015 | 30 |
| Katze | 0,015 | 85 |
Wie man sieht, haben die Wirbeltiere Frosch und Katze sehr dünne Axone, viel dünner als die des Tintenfischs. Trotzdem ist die Geschwindigkeit der Erregungsweiterleitung hier deutlich größer als beim Tintenfisch. Durch welchen "Trick" der Evolution ist diese beachtliche Steigerung der Geschwindigkeit möglich geworden?
Myelinscheiden isolieren das Axon
Bei Wirbeltieren liegen Axone von peripheren Nervenzellen (das sind Nervenzelle außerhalb des Gehirns) in der Regel nicht "nackt" vor, sondern sie sind von einer Markscheide umgeben. Das ist durchaus vergleichbar mit den Kupferkabeln in einem elektrischen Gerät, die von Kunststoff umgeben sind. Der Kunststoff um die Kabel hat die Aufgabe, die einzelnen Kabel voneinander zu isolieren, damit kein Kurzschluss entsteht. Eine ähnliche Aufgabe hat die Markscheide, die ein Axon umgibt. Würden sich zwei Axone direkt berühren, so könnten Aktionspotenziale des einen Axons zur Bildung von Aktionspotenzialen auf dem anderen Axon führen, wo diese vielleicht gar nicht erwünscht sind.

Hier sehen wir ein Axon, das von acht SCHWANNschen Zellen umgeben ist. Die Gesamtheit aller SCHWANNschen Zellen eines Axons wird als Markscheide oder - nach einem chemischen Bestandteil ihrer Zellmembranen - Myelinscheide bezeichnet. In den Lücken zwischen zwei SCHWANNschen Zellen ist das Axon gut zu sehen. Diese Lücken werden als RANVIERsche Schnürringe bezeichnet. Die SCHWANNschen Zellen gehören übrigens zu einer besonderen Gruppe von Zellen des Nervensystems, den sogenannten Gliazellen (Einzelheiten siehe dort).

Hier sehen wir einen Querschnitt durch ein Axon (gelb), das von einer SCHWANNschen Zelle umhüllt ist, deren Zellkern oben zu sehen ist.
In Wirklichkeit sind es noch mehr Schichten (zum Teil Hunderte), die ein Axon umhüllen, aber ich hatte keine Geduld mehr beim Zeichnen des Bildes und habe mich mit vier Schichten begnügt.
Myelinscheiden erhöhen die Geschwindigkeit der Erregungsleitung
Es ist kein Zufall, dass die Axone der Wirbeltiere so dünn sind und trotzdem die Erregung ungleich schneller weiterleiten als die teils sehr dicken Axone der Wirbellosen. Verantwortlich hierfür ist die Myelinscheide, die das Axon nicht nur isoliert, sondern auch für die hohe Geschwindigkeit der Erregungsleitung verantwortlich ist.
Bei einem marklosen Axon ist es ja so, dass ein Aktionspotenzial an einer bestimmten Stelle des Axons ein neues Aktionspotenzial in der benachbarten Region induziert. Dieses ruft dann wiederum ein Aktionspotenzial in der nächsten Region hervor und so weiter. Die so erzielte Geschwindigkeit ist schon recht beeindruckend, reicht aber für viele Zwecke nicht aus.
Bei einem myelinisierten Axon befinden sich die wichtigen spannungsgesteuerten Natrium-Kanäle nur in der Membran der RANVIERschen Schnürringe. Nur dort können Aktionspotenziale entstehen. Schauen wir uns nun mal an, wie das Membranpotenzial eines Axons mit vier SCHWANNschen Zellen aussieht:
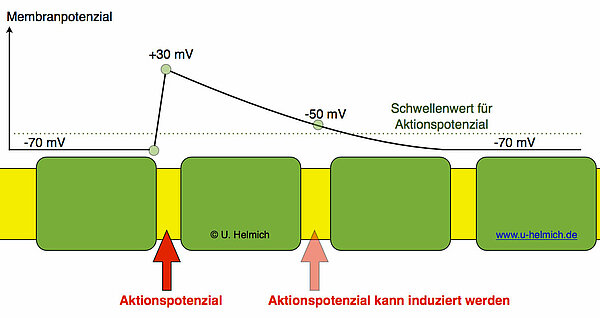
An dem Schnürring, an dem gerade ein Aktionspotenzial herrscht, hat das Membranpotenzial einen Wert von +30 mV. Am rechts benachbarten Schnürring kann man in diesem Augenblick ein Membranpotenzial von vielleicht -47 mV messen. Das Membranpotenzial liegt also deutlich über dem Wert im Ruhezustand. Diese überschwellige Depolarisierung am benachbarten Schnürring reicht zur Induktion eines neuen Aktionspotenzials völlig aus. Das Aktionspotenzial "spring" also quasi von dem einen Schnürring zum nächsten Schnürring. Die Zeitspanne für diesen "Sprung" ist sehr kurz, und wenn am benachbarten Schnürring das Aktionspotenzial entstanden ist, wird bereits am nächsten Schnürring ein neues Aktionspotenzial induziert. Wegen dieser "Sprünge" nennt man diese Art der Erregungsweiterleitung auch saltatorische Erregungsleitung (salto = Sprung).
Für Experten:
Manchmal liest man in Schulbüchern, dass im Innern des Axons Ausgleichströmchen stattfinden: Die bei einem Aktionspotenzial eingedrungenen Natrium-Ionen diffundieren nach links und rechts und sorgen so für eine überschwellige Depolarisierung bei den beiden benachbarten Schnürringen. Diese Diffusion durch Ausgleichströmchen mag ja auch eine gewisse Rolle bei der Induktion neuer Aktionspotenziale spielen, aber eigentlich kann man durch einfache Diffusionsvorgänge nicht die enormen Geschwindigkeiten erklären, die bei der saltatorischen Erregungsleitung erzielt werden. Vor allem dann nicht, wenn der Abstand zwischen den Schürringen 1mm oder mehr beträgt, wie es bei manchen großen Axonen der Fall ist.
Für richtige Experten:
Das Axon wird durch die SCHWANNschne Zellen extrem stark isoliert; auf der Außenseite des Axons befinden sich über 100 Lagen von Lipid-Doppelschichten. Für positive oder negative Ionen ist da kein Platz mehr. Das heißt wiederum, dass die Ionen, die sich auf der Innenseite der Axonmembran angesammelt haben, nicht von der entgegengesetzt geladenen Außenseite angezogen werden, wie das bei einem marklosen Axon der Fall ist (und bei den Schnürringen). Mit anderen Worten, die eingedrungenen Natrium-Ionen können sich auf der Innenseite des Axons völlig "ungebremst" ausbreiten, mit wesentlich größerer Geschwindigkeit als in einem marklosen Axon. Insofern kommt den Ausgleichströmchen doch eine größere Bedeutung zu. Siehe dazu auch die Vertiefungsseite.
Wirbeltierneuronen haben also die saltatorische Erregungsleitung "erfunden", um das Problem zu lösen, dass große Bündel aus vielen Axonen (= Nervenfasern) viele Impulse (= Aktionspotenziale) möglichst schnell leiten müssen, ohne dabei übermäßig dick sein zu müssen, wie das bei marklosen Neuronen der Fall wäre.
Myelinscheiden bestehen aus mehreren SCHWANNschen Zellen, die die Axone mehrlagig umwickeln und dabei elektrisch isolieren. Die RANVIERschen Schnürringe zwischen den SCHWANNschen Zellen ermöglichen die saltatorische Erregungsleitung: Ein Aktionspotenzial an einem Schnürring induziert ein neues Aktionspotenzial am nächsten Schnürring. Die dabei erzielte Geschwindigkeit ist selbst bei sehr dünnen Axonen sehr groß.